
角膜上皮是位于眼表最外層的非角質化復層磷狀上皮,作為屈光系統的屏障結構,對維持眼球內部環境穩定發揮了重要作用,其完整性和透明性并決定了視覺質量。目前研究表明,角膜和球結膜交界處的角膜緣區域存在著角膜緣干細胞(limbal stem cell,LSC),是角膜上皮更新和損傷修復的主要來源。位于角膜緣基底層的LSC不斷進行自我更新和分化,并向中央角膜遷移來替代死亡或受損的角膜上皮細胞(corneal epithelial cell,CEC),從而維持角膜上皮的穩態。在臨床上,LSC缺乏或病變會導致角膜混濁、新生血管侵入、上皮結膜化、角膜基質排列紊亂等癥狀,是世界上主要致盲原因之一。因而,探究LSC命運決定和穩態維持的機理具有重要意義。
哺乳動物成體干細胞的命運決定、自我更新和分化主要由染色質的表觀遺傳結構和轉錄調控網絡支配。染色質順式調控增強子元件,尤其是超級增強子,通過招募轉錄因子并與啟動子區域的轉錄復合物相互作用,從而驅動細胞或組織特異基因的表達。近年來,核心轉錄調控環路(core transcription regulatory circuitry,CRC)模型,即超級增強子調控的多個轉錄因子反過來相互調控各自的超級增強子,被證明在決定組織特異性和疾病發生過程中發揮了重要作用。大量研究表明,干細胞特性、可塑性和穩態由多個核心轉錄因子的協同作用通過建立特定的染色質表觀遺傳結構來實現。研究表明,PAX6作為LSC命運決定的master regulator,對角膜上皮發育起到必不可少的作用。然而,LSC的表觀遺傳圖譜及關鍵轉錄因子尚待進一步探究,LSC中的轉錄因子調控網絡也未有報道。
2021年1月18日,來自我校中山眼科中心的歐陽宏及劉奕志團隊在Nature communications雜志以research article形式發表題為“Core transcription regulatory circuitry orchestrates corneal epithelial homeostasis”的研究論文。該工作首次描繪了人類LSC的組蛋白修飾和染色質可及性圖譜,構建了基于超級增強子與轉錄因子互作的轉錄調控網絡。鑒定超級增強子基因RUNX1和SMAD3是LSC的命運決定和穩態維持必不可少的轉錄因子。發現RUNX1、PAX6和SMAD3(RPS)蛋白相互作用,并形成一個LSC特異的CRC。特別地,RUNX1促進了LSC中超級增強子和普通增強子區域的H3K27ac修飾但不影響H3K4me1、H3K4me3和H3K27me3。該工作也揭示,破壞RPS的協同作用與多種常見角膜疾病相關。
在這項研究工作中,研究人員通過解析LSC的4種組蛋白修飾 (H3K27ac, H3K4me1, H3K4me3, H3K27me3)和染色質可及性圖譜,構建了CRC調控網絡,并篩選出RUNX1和SMAD3作為潛在的候選關鍵因子(圖1)。
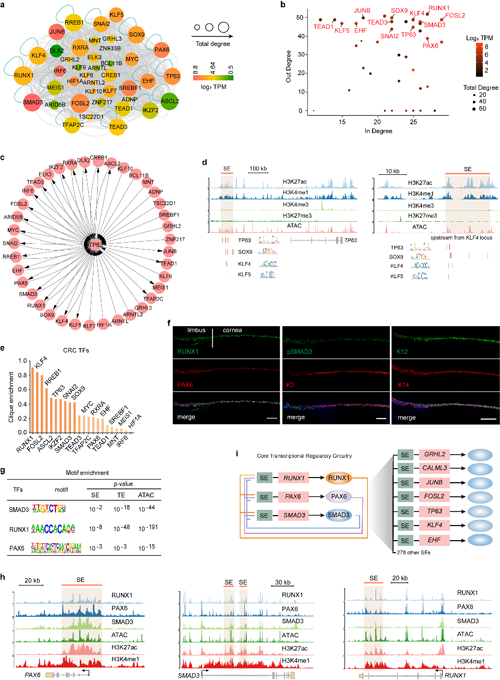
圖1 LSC特異的CRC轉錄因子調控網絡
利用ChIP-Seq分析表明,超級增強子調控的RPS反過來相互共調控各自的超級增強子,也共同調控了額外的超級增強子網絡,形成了一個LSC特異的CRC(圖1)。在LSC中分別敲低RUNX1和SMAD3誘導LSC特性基因包括PAX6的表達明顯下調,而角質化皮膚上皮特異基因的表達顯著上調。RUNX1或SMAD3敲低后的LSC會進一步分化為角質化的皮膚樣上皮細胞(圖2)。該文證明CRC轉錄因子RUNX1和SMAD3對維持LSC的命運、特性和可塑性發揮了必不可少的作用。
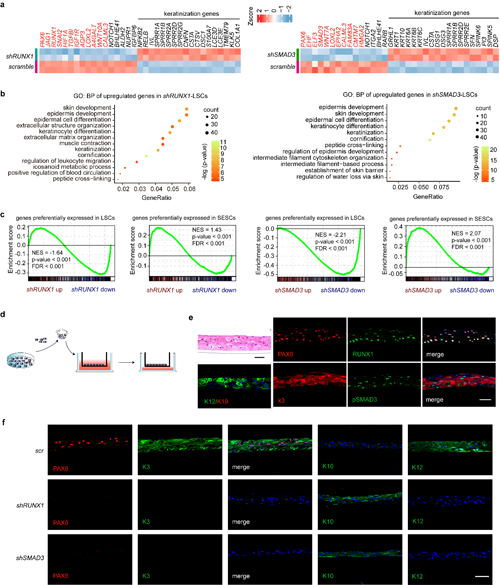
圖2 RUNX1或SMAD3敲降誘導LSC命運轉變
進一步的免疫共沉淀實驗顯示,RUNX1、PAX6和磷酸化SMAD3蛋白相互作用,形成三元復合物。通過聯合分析RNA-Seq和ChIP-Seq數據,發現RPS在基因組上共定位,通過協同調控超級增強子和普通增強子活性來影響維持LSC功能和特性關鍵基因的表達(圖3)。
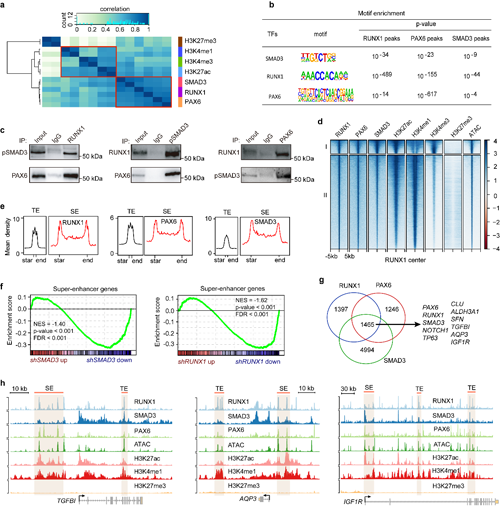
圖3 RPS復合物正向調控超級增強子活性
接下來,在LSC中比較對照組和RUNX1干擾組的組蛋白修飾,發現干擾RUNX1導致了下游關鍵基因的超級增強子或普通增強子的H3K27ac水平顯著降低,但對H3K4me1、H3K4me3和H3K27me3沒有影響。敲低RUNX1破壞了大量超級增強子的形成,導致超級增強子轉變為普通增強子(圖4),但干擾SMAD3對這4種組蛋白修飾均無影響。
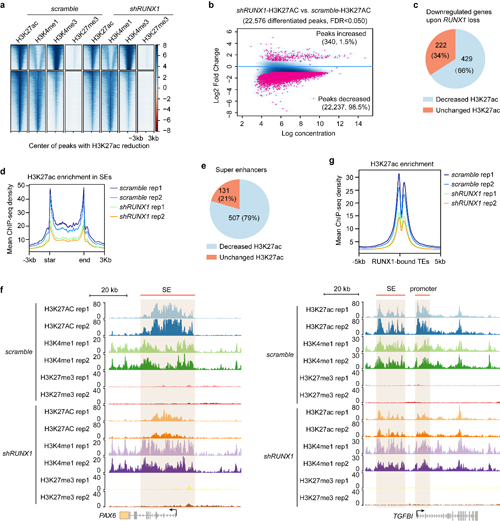
圖4 RUNX1敲降破壞LSC譜系特異增強子的H3K27ac修飾
最后,研究人員通過分析臨床上常見的幾種角膜疾病(炎癥、潰瘍、白斑、堿燒傷),發現在一些病變的角膜上皮區域,角膜上皮特性標記基因PAX6和KRT12的表達被沉默,轉而皮膚上皮標記基因KRT1和KRT10被激活。在這些角質化病變區域也觀察到RUNX1的缺失,但pSMAD3仍然被保留。pSMAD3由于沒有互作蛋白RUNX1和PAX6的協助,它的單獨存在不能維持角膜上皮特性,證明了RPS協同作用異常與人類角膜疾病發生密切相關(圖5)。
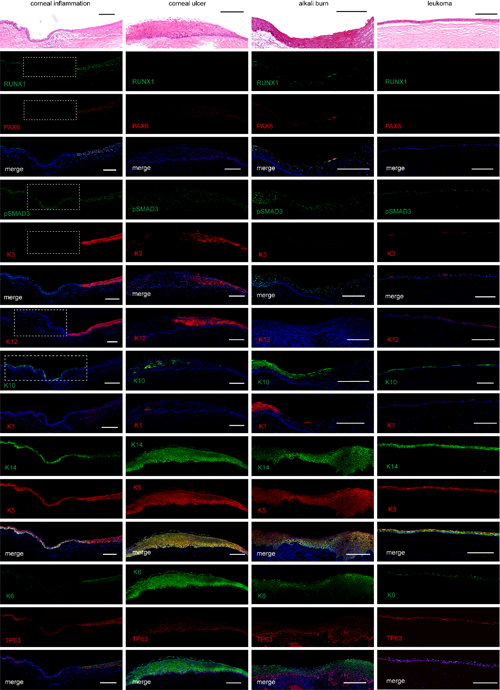
圖5 RPS與角膜上皮病理學改變
綜上,該研究首次描繪了人LSC的染色質景觀,鑒定RUNX1和SMAD3作為LSC的核心轉錄因子,與PAX6形成CRC通過協同調控超級增強子來維持角膜上皮的特性和穩態。本研究揭示了決定成體干細胞命運、特性和可塑性的轉錄因子協同作用網絡(圖6)。
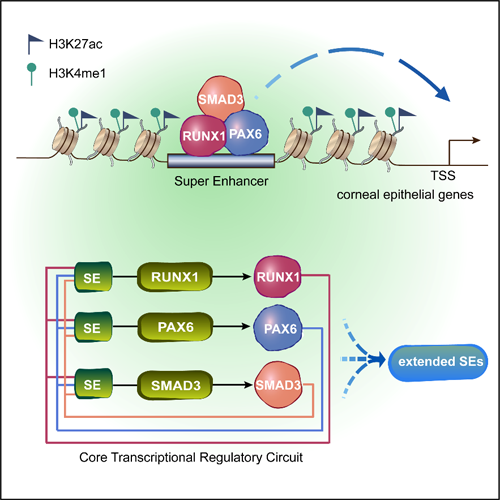
圖6 譜系特異轉錄因子建立的CRC模型決定角膜上皮特性


