An important progress on the mechanisms for global plant distribution was made by Prof. Shaolin Peng’s team at the School of Life Sciences
Source: School of Life Sciences
Edited by: Tan Rongyu, Wang Dongmei
CSR-strategic scheme was proposed by the famous UK ecologist, Dr. Philip Grime, to describe the triangular trade-off between three important plant functional strategies, competitor (C), stress tolerator (S) and ruderal (R). These three strategies may benefit plant range expansion under different circumstances. However, how the three strategies influence the extent of plant global distribution remains to be explored. The recent study entitled “The role of functional strategies in global plant distribution” by Prof. Shaolin Peng’s team at the School of Life Sciences, Sun Yat-sen University provides an answer to this important question (Fig. 1a) and directly compared the difference in the dominant functional strategies influencing plant distributions in the native and naturalized ranges (Fig. 1b).
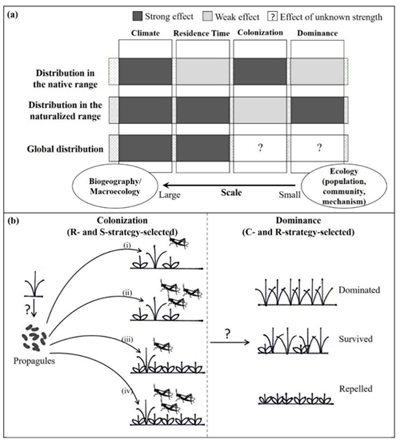
Fig. 1. The theoretical background for the potentially different dominant functional strategies dictating the native, naturalized, and global distributions of plant species. (a) The effects of major ecological factors on the species distribution in the native, naturalized, and global distributional ranges. The question marks denote the areas that have not been tested yet. (b) The colonization and dominance processes associated with the ruderal (R), competitor (C) and stress-tolerator (S) strategies. The question marks denote the processes whose outcomes depend on the functional strategies of the focal species.
In this study, a dataset of 2645 vascular plant species was compiled to explore the roles of the CSR strategies in global plant distribution. The functional strategy-range size associations differed across different distributional ranges and growth forms. Specifically, species global and native range sizes were positively associated with the R score; species naturalized range size was positively associated with the C score; and all range-size measurements were negatively associated with the S score (Fig. 2). These patterns were mostly driven by herbs but not shrubs or trees. For species global and native-range distributions, the patterns of shrubs were even opposite to those of herbs (Fig. 2). These findings suggest that although the native range size is a strong indicator of the invasion success of a species, we may overestimate the invasion potential of ruderal trees and herbs solely based on their native range sizes because the R strategy has a greater advantage in the native range than in the naturalized range for these species. Along the same lines, we may underestimate the invasion potential of the C-selected native herbs, because the C strategy has a greater advantage in the naturalized range than in the native range.
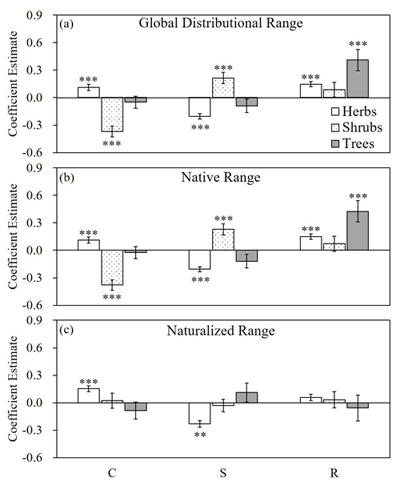
Fig. 2. Effect of the C/S/R score on the three range-size measurements, (a) global range size, (b) native range size and (c) naturalized range size, for each growth form based on the phylogenetic generalized least square (PGLS) models. Estimates for the effects with standard errors are shown. The estimates that are significantly different from zero are marked by asterisks (* P≤0.05; ** P≤0.01; and *** P≤0.001).
More importantly, these findings emphasize the importance of distinguishing the functional strategies associated with native and non-native species to provide better guidance for nature conservation. Because narrow-ranging native plants usually possess a weak R strategy (i.e., a weak dispersal ability), there is a high possibility of conserving these species by increasing propagule pressures and aiding dispersal to distant habitats. Meanwhile, because non-native herbs suggest that the wide-ranging (highly invasive) herbs in the naturalized range usually possess a strong C strategy and a weak S strategy (i.e., a strong competitiveness and a weak stress tolerance), there is a high possibility of controlling these species by enhancing the resistance of resident community via plant diversity manipulation and restrict disturbances that will alleviate resource limitation in natural communities.
This work was published in
Ecography (IF=6.455, JCR=Q1), which was supported by the National Natural Science Foundation of China, the Natural Science Foundation of Guangdong Province (China) and the Guangdong Provincial Key Laboratory of Plant Resources (China). Prof. Shaolin Peng is the corresponding author, and Dr. Huixuan Liao is the first author of this paper.
Link to the paper:
https://onlinelibrary.wiley.com/doi/10.1111/ecog.05476







